Нервно-мышечные веретена представляют собой рецепторы растяжения, расположенные в пределах произвольной поперечно-полосатой мускулатуры; благодаря своей активности они способны фиксировать состояние растяжения мышц и отправлять собранную информацию в спинной мозг и в головной мозг. Поэтому активность нервно-мышечных веретен очень важна как для предотвращения травм, связанных с чрезмерным удлинением, так и для поддержания нормального мышечного тонуса, а также для выполнения плавных и контролируемых движений жидкости.
Все скелетные мышцы, за исключением нижней челюсти, содержат в себе различные нервно-мышечные веретена, которые особенно сконцентрированы на уровне жевательных мышц, позвоночника, глаз, конечностей и рук. Здесь нервно-мышечные веретена длиной около 5-10 мм расположены параллельно с обычными мышечными волокнами, и благодаря этому особому расположению «бок о бок» они способны фиксировать степень удлинения.
анатомия
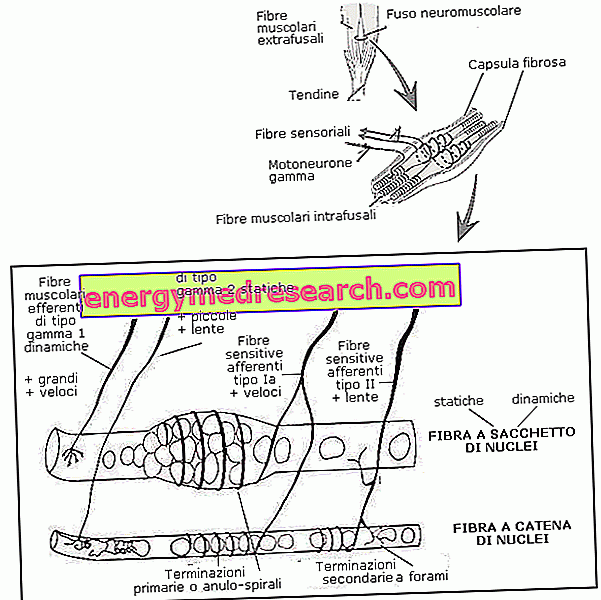
Нервно-мышечный веретено состоит из капсулы соединительной ткани, которая обволакивает небольшую группу мышечных волокон (от 4 до 10), снабженную «особой» цитологической структурой; эти волокна часто называют внутрифузальными, чтобы отличать их от обычных волокон, которым для ровного игрового поля дают прилагательное «extrafusali».
Физиология внутрифузальных волокон объясняется, прежде всего, детальным изучением анатомической структуры. В своих крайностях они очень похожи на обычные волокна и содержат для этого сократительные полосатые фибриллы. Реальная разница заключается в экваториальной части, которая кажется увеличенной, лишенной миофибрилл и богатой чувствительными к растяжению сенсорными окончаниями, погруженными в желатиновое вещество.
Поэтому говорят, что волокна нервно-мышечных веретен эффекторны к двум полюсам (они сжимаются в ответ на нервный раздражитель) и излучателям в центре (откуда они посылают информацию о состоянии удлинения).
С анатомической точки зрения, внутрифузальные мышечные волокна делятся на ядерные волокна мешка (также называемые волокнами мешка или сумки) и волокна ядерной цепочки. Первые имеют расширенную центральную область, богатую ядрами. С другой стороны, волокна ядерных цепей имеют удлиненное ядерное распределение, всегда сконцентрированное в экваториальной области, но также вытянутое на периферии; они также короче и тоньше предыдущих.
С анатомической точки зрения чувствительные окончания нервно-мышечного веретена располагаются, частично путем сворачивания в срединную область (анулярно-спиральные или первичные окончания) и частично образуя ветвь саженца в соседних областях (цветочные или вторичные окончания).
Первичные окончания более толстые, имеют высокую скорость проводимости, относятся к классу волокон Ia и отходят от волокон мешка и ядерных цепей; вторичные окончания, принадлежащие к классу волокон типа II, вместо этого более тонкие, менее быстрые при распространении импульсов и иннервируют в основном цепочечные волокна ядер.
С физиологической точки зрения, с другой стороны, мы различаем чувствительные к быстрой проводимости волокна (тип Ia) и чувствительные к медленной проводимости волокна (тип II). Первые, несмотря на наличие окончаний на обоих типах волокна, представляют собой концевые спирально-кольцевые окончания, характерные для волокон с волокнистым мешком динамических ядер (см. Ниже). Более медленные волокна II, с другой стороны, имеют концевые спирально-кольцевые окончания, которые обертывают волокна мешкового волокна из статических ядер и цепные волокна; также цветущие окончания принадлежат этой категории.
В отличие от экстрафузальных мышечных волокон, которые получают афференты от альфа-моторных нейронов, веретенообразные волокна сокращаются под действием гамма-мотонейронов (нервные волокна, идущие от переднего рога спинного мозга, характеризуются небольшим калибром).
ПРОДОЛЖЕНИЕ: Физиология нервно-мышечных веретен »