Чтобы иметь возможность говорить всесторонне о гемоглобине (Hb), полезно сначала разобраться с миоглобином (Mb), который очень похож на гемоглобин, но намного проще. Существуют строгие родственные связи между гемоглобином и миоглобином: оба являются конъюгированными белками, а их простетическая группа (небелковая часть) является гемовой группой.
Миоглобин представляет собой глобулярный белок, состоящий из одной цепи из примерно ста пятидесяти аминокислот (в зависимости от организма), а его молекулярный вес составляет около 18 кДа.
Как уже упоминалось, он снабжен гемовой группой, которая вставлена в гидрофобную (или липофильную) часть белка, состоящую из складок, связанных с α-спиральными структурами волокнистых белков.
Миоглобин состоит в основном из сегментов α-спиралей, присутствующих в количестве восьми, и состоит, почти исключительно, из неполярных остатков (лейцин, валин, метионин и фенилаланин), тогда как полярные остатки практически отсутствуют (аспарагиновая кислота, глутаминовая кислота, лизин и аргинин); единственными полярными остатками являются два гистидина, которые играют фундаментальную роль в атаке кислорода на гемовую группу.
Группа гемов представляет собой группу хромофоров (поглощает в видимой области) и является функциональной группой миоглобина.
Смотрите также: гликированный гемоглобин - гемоглобин в моче
Немного химии
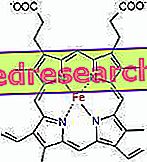
Связь между протопорфирином и железом является типичной связью так называемых координационных соединений, которые представляют собой химические соединения, в которых центральный атом (или ион) образует связи с другими химическими веществами в количестве, превышающем его степень окисления (электрический заряд). В случае гема эти связи являются обратимыми и слабыми.
Координационное число (число координационных связей) железа равно шести: вокруг железа может быть шесть молекул, которые разделяют связывающие электроны.
Чтобы сформировать координационное соединение, нужны две орбитали с правильной ориентацией: одна способна «покупать» электроны, а другая - жертвовать их.
В геме железо образует четыре плоские связи с четырьмя атомами азота, которые находятся в центре прото-порфиринового кольца, и пятую связь с проксимальным гистидиновым азотом; железо имеет шестую свободную координационную связь и может связываться с кислородом.
Когда железо находится в форме свободного иона, все его орбитали d- типа имеют одинаковую энергию; в миоглобине ион железа связан с протопорфирином и гистидином: эти виды магнитно нарушают орбитали железа; Степень возмущения будет различной для разных орбиталей в зависимости от их пространственной ориентации и ориентации нарушающих видов. Поскольку полная энергия орбиталей должна вернуться к постоянной, возмущение вызывает энергетическое разделение между различными орбиталями: энергия, полученная одними орбиталями, эквивалентна энергии, потерянной другими.
Если расстояние между орбиталями не очень велико, предпочтительна электронная схема с высоким спином: связывающие электроны пытаются расположить себя в параллельных спинах на нескольких возможных подуровнях (максимальная кратность); с другой стороны, если возмущение очень сильное и между орбиталями имеется большое расстояние, может оказаться более удобным соединить связывающие электроны на орбитали с более низкой энергией (низкий спин).
Когда железо связывается с кислородом, молекула приобретает низкоспиновое расположение, тогда как когда железо имеет шестую свободную координационную связь, молекула имеет высокий спин.
Благодаря этой разнице в спине, благодаря спектральному анализу миоглобина, мы можем понять, есть ли связанный кислород (MbO2) или нет (Mb).
Миоглобин является белком, типичным для мышц (но не только в мышцах).
Миоглобин извлекается из кашалота, в котором он присутствует в больших количествах, а затем очищается.
У китообразных дыхание подобно дыханию людей: имея легкие, они должны поглощать воздух через дыхательные процессы; кашалот должен нести как можно больше кислорода в мышцах, которые способны накапливать кислород, связывая его с присутствующим в них миоглобином; затем кислород медленно выделяется при погружении китообразного, потому что для его метаболизма требуется кислород: чем больше кислорода может поглотить кашалот, тем больше кислорода доступно во время погружения.

Чтобы сделать мясо более или менее красным, необходимо содержание гемопротеинов (именно гем делает мясо красным).
Гемоглобин имеет много структурных аналогий с миоглобином и способен обратимо связывать молекулярный кислород; но, в то время как миоглобин ограничен мышцами и периферическими тканями в целом, гемоглобин обнаружен в эритроцитах или эритроцитах (это псевдо-клетки, то есть они не являются настоящими клетками), которые составляют 40% крови.
Гемоглобин - это тетраметр, то есть он состоит из четырех полипептидных цепей, каждая с гемовой группой и две на две (у человека есть две альфа-цепи и две бета-цепи).
Основная функция гемоглобина - транспорт кислорода ; Еще одна функция крови, в которой участвует гемоглобин, - это транспортировка веществ в ткани.
На пути от легких (богатых кислородом) к тканям гемоглобин переносит кислород (в то же время другие вещества также достигают тканей), тогда как на обратном пути он несет с собой отходы, собираемые из тканей, особенно углекислый газ, образующийся в процессе обмена веществ.
В развитии человека существуют гены, которые экспрессируются только в течение определенного периода времени; По этой причине у нас разные гемоглобины: эмбриональные, эмбриональные, взрослого мужчины.
Цепочки, из которых состоят эти разные гемоглобины, имеют разную структуру, но схожесть функций, которые они выполняют, более или менее одинакова.
Объяснение присутствия нескольких различных цепей заключается в следующем: в процессе эволюции организмов также развился гемоглобин, специализирующийся на транспортировке кислорода из областей, богатых дефицитными областями. В начале эволюционной цепи гемоглобин переносил кислород в небольших организмах; в ходе эволюции организмы достигли больших размеров, поэтому гемоглобин изменился, чтобы иметь возможность транспортировать кислород в области, более отдаленные от точки, в которой он был богат; для этого в ходе эволюционного процесса были кодифицированы новые структуры цепей, составляющих гемоглобин.
Миоглобин связывает кислород даже при умеренном давлении; в периферических тканях давление (PO2) составляет около 30 мм рт. ст.: при этом давлении миоглобин не выделяет кислород, поэтому он неэффективен в качестве переносчика кислорода. Гемоглобин, с другой стороны, обладает более эластичным поведением: он связывает кислород при высоких давлениях и высвобождает его при снижении давления.
Когда белок функционально активен, он может слегка изменить свою форму; например, оксигенированный миоглобин имеет форму, отличную от не оксигенированного миоглобина, и эта мутация не влияет на соседние.
Дискурс отличается в случае ассоциированных белков, таких как гемоглобин: когда оксигенируется цепь, она вынуждена изменять свою форму, но эта модификация является трехмерной, поэтому другие цепочки тетраметра также затрагиваются. Тот факт, что цепи связаны друг с другом, заставляет думать, что модификация одной затрагивает другие соседние, даже если в разной степени; когда цепь оксигенирована, другие цепи тетраметра принимают «менее враждебное отношение» к кислороду: сложность, с которой оксигенируется цепь, уменьшается по мере того, как соседние цепи оксигенируют себя. Тот же аргумент действителен для дезоксигенации.
Четвертичная структура дезоксигемоглобина называется Т (напряженной) формой, а оксигемоглобина - формой R (высвобождается); в растянутом состоянии существует ряд довольно сильных электростатических взаимодействий между кислотными аминокислотами и основными аминокислотами, которые приводят к жесткой структуре дезоксигемоглобина (именно поэтому «напряженная форма»), тогда как когда кислород связан, сущность этих уменьшается взаимодействие (вот почему "высвобождается форма"). Кроме того, в отсутствие кислорода заряд гистидина (см. Структуру) стабилизируется противоположным зарядом аспарагиновой кислоты, в то время как в присутствии кислорода наблюдается тенденция потери белка протоном; Все это означает, что оксигенированный гемоглобин является более сильной кислотой, чем деоксигенированная гемоглобия: эффект Бора .
В зависимости от pH, гемовая группа более или менее легко связывается с кислородом: в кислой среде гемоглобин легче выделяет кислород (напряженная форма стабильна), в то время как в основной среде связь с кислородом сильнее.

Каждый гемоглобин выделяет 0, 7 протона на каждый моль кислорода (O2).
Эффект Бора позволяет гемоглобину улучшить его способность переносить кислород.
Гемоглобин, который совершает путешествие из легких в ткани, должен уравновешиваться в соответствии с давлением, pH и температурой.
Мы видим влияние температуры .
Температура в легочных альвеолах примерно на 1-1, 5 ° С ниже, чем наружная температура, в то время как в мышцах температура составляет около 36, 5-37 ° С; с ростом температуры коэффициент насыщения уменьшается (при одном и том же давлении): это происходит потому, что кинетическая энергия увеличивается и диссоциация благоприятна.

Существуют и другие факторы, которые могут влиять на способность гемоглобина связываться с кислородом, одним из которых является концентрация 2, 3-бисфосфоглицерата .
2, 3-бисфосфоглицерат представляет собой метаболический агент, присутствующий в эритроцитах в концентрации 4-5 мМ (ни в одной другой части тела он не присутствует в такой высокой концентрации).
При физиологическом pH 2, 3-бисфосфоглицерат депротонируется и имеет пять отрицательных зарядов; он вклинивается между двумя бета-цепями гемоглобина, потому что эти цепи имеют высокую концентрацию положительных зарядов. Электростатические взаимодействия между бета-цепями и бисфосфоглицератными цепями придают системе определенную жесткость: получается напряженная структура, которая имеет небольшое сродство к кислороду; во время оксигенации 2, 3-бисфосфоглицерат удаляется.
В эритроцитах есть специальный аппарат, который превращает 1, 3-бисфосфоглицерат (образующийся в результате метаболизма) в 2, 3-бисфосфоглицерат, так что он достигает концентрации 4-5 мМ и, следовательно, гемоглобин способен обменивать l кислород в тканях.
Гемоглобин, поступающий в ткань, находится в высвобожденном состоянии (связан с кислородом), но в непосредственной близости от ткани он карбоксилируется и переходит в растянутое состояние: белок в этом состоянии имеет более низкую тенденцию связываться с кислородом, по сравнению с освобожденным состоянием, поэтому гемоглобин выделяет кислород в ткани; кроме того, из-за реакции между водой и диоксидом углерода происходит образование ионов H +, а следовательно, и кислорода из-за эффекта Бора.
Углекислый газ диффундирует в эритроцит, проходя через плазматическую мембрану; учитывая, что эритроциты составляют около 40% крови, следует ожидать, что только 40% углекислого газа, который диффундирует из тканей, поступает в них, в действительности 90% углекислого газа попадает в эритроциты, потому что они содержат фермент, который превращает Из диоксида углерода в углекислоте следует, что стационарная концентрация диоксида углерода в эритроцитах низкая и, следовательно, скорость проникновения высока.

Другое явление, которое возникает, когда эритроцит достигает ткани, заключается в следующем: по градиенту HCO3- (производное углекислого газа) выходит из эритроцита, и, чтобы уравновесить выход отрицательного заряда, мы имеем поступление хлоридов, которое определяет повышение осмотического давления: для уравновешивания этого изменения также имеется попадание воды, которая вызывает набухание эритроцитов (эффект ГАМБУРГЕРА). Противоположное явление происходит, когда эритроцит достигает легочных альвеол: происходит дефляция эритроцитов (эффект HALDANE). Таким образом, венозные эритроциты (прямые к легким) более круглые, чем артериальные.


