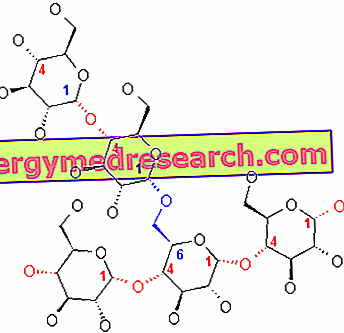
Гликоген является макромолекулой (молекулярная масса около 400 миллионов дальтон) α-глюкозы, в которой в основном присутствуют α-1, 4-гликозидные связи и ответвления в соотношении 1:10, благодаря α-1, 6-гликозидным связям.
Гликоген является резервным материалом и постоянно разлагается и восстанавливается; во всей клеточной массе тела мы имеем около 100 г гликогена: большая его часть находится в печени, где он подвижен, и, следовательно, может использоваться как резерв для других органов (гликоген в мышцах не подвижен).
Ферменты, которые катализируют деградацию и синтез гликогена, все находятся в цитоплазме, поэтому необходима система регуляции, которая делает один путь неактивным, когда другой активен: если глюкоза доступна, последний превращается в гликоген (анаболизм), который является резервом, и наоборот, если требуется глюкоза, то гликоген разлагается (катаболизм).
Фермент, в основном участвующий в деградации гликогена, представляет собой гликогенфосфорилазу ; этот фермент способен расщеплять α-1, 4-гликозидную связь, используя в качестве литического агента неорганический ортофосфат: расщепление происходит посредством фосфорилирования и получается глюкозо-1-фосфат.
В пяти или шести единицах от точки разветвления фермент гликогенфосфорилаза больше не способен действовать, поэтому он отделяется от гликогена и заменяется разветвляющимся ферментом, который представляет собой трансферазу : в каталитическом сайте этого фермента имеется 'гистидин, который позволяет переносить три сахаридных звена к ближайшей гликозидной цепи (гистидин атакует первый углерод молекулы глюкозы). Только что упомянутый фермент представляет собой гликозилтрансферазу ; в конце действия этого фермента на боковой цепи остается только одна единица глюкозы, причем первый углерод связан с шестым углеродом глюкозы в основной цепи. Последняя глюкозная единица боковой цепи высвобождается под действием фермента α-1, 6-гликозидазы (этот фермент является второй частью расщепляющего фермента); учитывая, что ответвления в гликогене находятся в соотношении 1:10, из полной деградации макромолекулы мы получаем около 90% глюкозо-1-фосфата и около 10% глюкозы.
Действие вышеупомянутых ферментов позволяет устранить боковую цепь из молекулы гликогена; активность этих ферментов может повторяться до тех пор, пока не произойдет полная деградация цепи.
Давайте рассмотрим гепатоцит; глюкоза (усваивается через диету), когда она попадает в клетку, она превращается в 6-фосфат глюкозы и, таким образом, активируется. Глюкозо-6-фосфат под действием фосфоглюкомутазы превращается в глюкозо-1-фосфат: последний является непосредственным предшественником биосинтеза; в биосинтезе используется активированная форма сахаров, которая представлена сахаром, связанным с дифосфатом: обычно уридилдипосфатом (UDP). 1-фосфат глюкозы затем превращается в UDP-глюкозу; Этот метаболит находится под действием гликогенсинтазы, которая способна связывать UDP-глюкозу с невосстанавливающим концом растущего гликогена: удлиненный гликоген получают из глюкозидной единицы и UDP. UDP превращается ферментом нуклеозидифосфокиназой в UTP, который возвращается в кровообращение.
Разложение гликогена происходит под действием гликогенфосфорилазы, которая высвобождает молекулу глюкозы и превращает ее в 1-фосфат глюкозы. Затем фосфоглюкомутаза превращает глюкозо-1-фосфат в глюкозо-6-фосфат.
Гликоген синтезируется, прежде всего, в печени и мышцах: в организме содержится 1-1, 2 грамма гликогена, распределенного по всей мышечной массе.
Гликоген миоцита представляет собой запас энергии только для этой клетки, в то время как гликоген, содержащийся в печени, также является резервом для других тканей, то есть его можно отправлять в виде глюкозы в другие клетки.
Глюкозо-6-фосфат, полученный в мышцах в результате разложения гликогена, затем направляется, в случае потребности в энергии, в гликолиз; в печени глюкозо-6-фосфат превращается в глюкозу под действием глюкозо-6-фосфат-фосфатазы (фермента, характерного для гепатоцитов) и попадает в кровоток.
Как гликогенсинтаза, так и гликогенфосфорилаза воздействуют на невосстанавливающие единицы гликогена, поэтому должен быть гормональный сигнал, который управляет активацией одного пути и блокированием другого (или наоборот).
В лаборатории удалось удлинить гликогеновую цепь, используя гликогенфосфорилазу и используя глюкозо-1-фосфат в очень высокой концентрации.
В клетках гликогенфосфорилаза только катализирует реакцию разложения, потому что концентрации метаболитов таковы, что сдвигают баланс следующей реакции вправо (то есть в сторону разложения гликогена):

Давайте рассмотрим механизм действия гликогенфосфорилазы: уксусный кислород (который действует как мостик между глюкозными единицами) связывается с фосфорильным водородом: промежуточный продукт реакции образуется карбокатионом (на глюкозе, которая является конечности), с которым фосфорил (Pi) связывается очень быстро.
Гликогенфосфорилаза требует кофактора, который представляет собой пиридоксальфосфат (эта молекула также является кофактором для трансаминаз): фосфорил протонирован только частично (пиридоксальфосфат окружен гидрофобной средой, что оправдывает присутствие протонов, связанных с ней), Фосфорил (Pi) способен превращать протон в гликоген, потому что такой фосфорил восстанавливает протон из частично протонированного фосфорила пиридоксальфосфата. Вероятность того, что при физиологическом pH фосфорил теряет протон и остается полностью депротонированным, очень мала.
Теперь посмотрим, как работает фосфоглюкомутаза. Этот фермент имеет фосфорилированный сериновый остаток в каталитическом сайте; серин превращает фосфорил в глюкозо-1-фосфат (в шестом положении): в течение короткого времени образуется 1, 6-бисфосфат глюкозы, затем серин перефосфорилируется, принимая фосфорил в первом положении. Фосфоглюкозная мутаза может работать в обоих направлениях, то есть превращать глюкозо-1-фосфат в глюкозо-6-фосфат или наоборот; если образуется глюкозо-6-фосфат, его можно направить непосредственно в гликолиз, в мышцы или преобразовать в глюкозу в печени.
Фермент уридилфосфоглюкстрансфераза (или UDP-глюкоза-пирофосфорилаза) катализирует реакцию переноса глюкозо-1-фосфата в UTP посредством фосфорильной атаки a.
Только что описанный фермент представляет собой пирофосфорилазу: это название связано с тем, что реакция, противоположная только что описанной, представляет собой пирофосфорилирование.
UDP глюкоза, полученная, как описано, способна удлинить гликогенную цепь моносахаридного звена.
Можно развить реакцию в направлении образования UDP-глюкозы, удаляя продукт, который является пирофосфатом; фермент пирофосфатаза превращает пирофосфат в две молекулы ортофосфата (гидролиз ангидрида) и, тем самым, поддерживает концентрацию пирофосфата настолько низкой, что процесс образования глюкозы UDP ускоряется термодинамически.
Как уже упоминалось, UDP глюкоза, благодаря действию гликогенсинтазы, способна удлинять гликогеновую цепь.
Последствия (в соотношении 1:10) связаны с тем, что, когда цепь гликогена состоит из 20-25 единиц, вмешивается разветвляющийся фермент (с гистидином на его каталитическом сайте), способный переносить серию из 7 -8 гликозидных единиц дальше по ходу 5-6 единиц: таким образом генерируется новое ветвление.
По причинам нервного происхождения или если энергия необходима из-за физических усилий, адреналин выделяется из надпочечников.
Клетками-мишенями адреналина (и норэпинефрина) являются клетки печени, мышц и жировой ткани (в последних происходит деградация триглицеридов и циркуляция жирных кислот: поэтому в митохондриях вырабатывается глюкоза 6). -фосфат, предназначенный для гликолиза, в то время как в адипоцитах глюкозо-6-фосфат под действием фермента глюкозо-6-фосфат-фосфатазы превращается в глюкозу и экспортируется в ткани).
Посмотрим теперь, как действует адреналин. Адреналин связывается с рецептором, расположенным на клеточной мембране (миоцитов и гепатоцитов), и это определяет трансляцию сигнала снаружи внутрь клетки. Активируется протеинкиназа, которая действует одновременно на системы, которые регулируют синтез и деградацию гликогена:
Гликогенсинтаза существует в двух формах: дефосфорилированная форма (активная) и фосфорилированная форма (неактивная); протеинкиназа фосфорилирует гликогенсинтазу и блокирует ее действие.
Гликогенфосфорилаза может существовать в двух формах: активная форма, в которой присутствует фосфорилированный серин, и неактивная форма, в которой серин дефосфорилирован. Гликогенфосфорилаза может быть активирована ферментом гликогенфосфорилазакиназа . Гликогенфосфорилазная киназа активна, если она фосфорилирована, и неактивна, если она дефосфорилирована; протеинкиназа имеет в качестве субстрата гликогенфосфорилазную киназу, то есть она способна фосфорилировать (и, следовательно, активировать) последнюю, которая, в свою очередь, активирует гликогенфосфорилазу.
Как только сигнал адреналина закончится, эффект, который он оказывает в клетке, также должен прекратиться: затем ферменты фосфатазы вмешиваются в белковые виды.